Fertility and Disease
The discovery the NORTIA is an MLO gene led to the hypothesis that pollen tubes and fungal hyphae share common mechanisms for establishing communication with a "host" cell. In both cases, a tip-growing cell makes contact with another cell and induces the polar localization of an MLO protein to the site of entry (Bhat et al., 2005; Kessler et al., 2010). In nta mutant ovules, the PT enters the synergid but does not succeed in communicating with the synergid and therefore does not rupture and deliver the sperm. In mlo mutants, fungal hyphae do not succeed in establishing a successful infection and therefore cannot reproduce (Consonni et al., 2006). Since fer and nta share a common phenotype in PT reception and FER is also expressed in vegetative tissues, we examined fer mutants for PM resistance and found a similar resistance phenotype to that seen in mlo mutants (Kessler et al., 2010). This data led to the intriguing hypothesis that FER acts as a sensor of extracellular perturbations and cooperates with MLOs to allow signaling with and entry of tip-growing cells (Kessler et al., 2010). This model is supported by the result that NTA redistribution upon PT arrival requires active FER (Kessler et al., 2010), but determining whether this is a direct or indirect pathway requires more investigation.
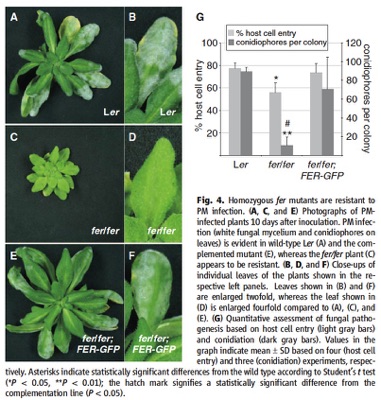
feronia mutants are powdery mildew resistant. (figure from Kessler, et al., 2010)
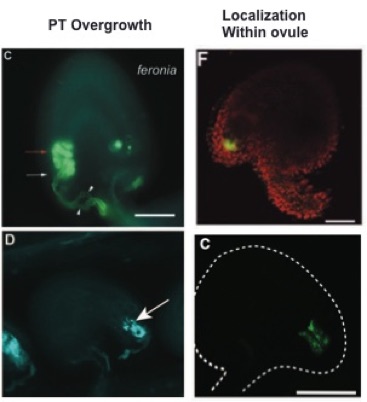
feronia (C) and nortia (D) mutants have similar pollen tube overgrowth phenotyps. (Figure adapted from Escobar, et al., 2007 and Kessler, et al., 2010)